Last week I attended the Annual Meetings of the (other) ESA in Austin, TX. Actually, I kinda helped organize that meeting. (More about that in my next post, which will focus on my time on the Program Committee.)
When I first submitted the title for the poster in June I had only just started this blog and I thought it would be a great idea to cover the numerous posts I would have written by November.
So, yeah, about that….
Still, it was a great exercise to go through and it helped keep me sane during the weeks leading up to the meeting.
Below is a picture of what it looked like at the conference. I was unable to spend much time with it since the official social hour for the poster session was right at the time of various committee meetings (obviously poor planning on the part of the Program Committee). Judging by the bump in views at my blog some people did find it interesting.
As I mention on the poster, social media has enriched my scientific life. One of the best parts of the meeting was therefore to meet, or catch-up, with some of my ento-tweeps. (Bummed that I had to miss the “official” tweetup).
One of my major “accomplishments” as Program Co-Chair was to get ESA to provide these twitter stickers for name-badges:
For more details on why twitter (and, in my opinion, other social media outlets) can be useful to entomologists please read @derekhennen‘s take at EntomologyToday.
To end this short post I’ll just include a tweet from @bug_girl because it reflects my sentiments exactly! Ento Bloggers Rule!
I traveled extensively over the summer; to Austin, TX, all through Western Europe and back to Illinois. All the while I was working on this blog post about bioinspired air-conditioning, which was appropriate because everywhere I went I seemed to have to suffer through heat-wave after heat-wave.
While wishing for Europe to have more air-conditioning units (especially in class-rooms and lecture halls), one of course wonders if that would only exacerbate the problem and make summers even hotter. Progress is constantly being made on making air-condition units more compact, more energy efficient, and thus more environmentally friendly. Inspiration on how to accomplish this has been already been found in natural systems.
Recently Brian Clark Howard wrote an interesting and popular article for National Geographic entitled: “5 Natural Air-Conditioning Designs Inspired by Nature”. Arthropods (termites & ticks) were prominently represented on this list. However, I would like to provide a little bit more detail and corrections to the NG’s list.
The insect examples touched upon in the NG article is that of the termite mound. The most famous architectural example of biomimicry or bioinspiration is the Eastgate Centre Building in Harare, Zimbabwe, which opened in 1996. Architect Mick Pearce and the firm Arup were supposedly partly inspired to build a building suitable for a tropical climate by considering the locally present termite mounds, and build their vision using locally available materials.
Eastgate Centre (with chimneys on roof) in Harare Zimbabwe. (Source: wikipedia.org. Unknown photographer)
Anyone who is even the slightest bit interested in biomimicry knows of the Eastgate Centre, but since the political and economic situation in Zimbabwe has probably only deteriorated since 1996 I was wondering if it was actually still standing and still used as the commercial center it had been envisioned as. So I took my questions to Twitter: [View the story “EastGate Building Harare, Zimbabwe” on Storify]. Turns out that the Eastgate building is still used and stands out prominently in the heart of Harare (and as @ardeans pointed out the building is located right on Robert Mugabe Road, sigh). One of the occupants is actually the United States Embassy.
It is just kind of too bad that the building is based on incorrect biology. Or is it?
First a little bit of background about the inspirational insect. Macrotermes termites (Macrotermitinae) occur over tropical Africa and Asia. There are about 330 species in this genus of relatively large termites. Most of the species build elaborate mounds. The tallest mounds occur in Africa (max of 30 feet, 9 meters). Macrotermes termites cultivate fungi, and spend most of their time, somewhere deep within the mound. The Macrotermes species that has been studied the most is the African species M. michaelseni.
Popular wisdom says that in order to optimize fungus growth the interior of the mound needs to be maintained within the narrow range of 29-32°C. To keep keep within this narrow temperature range, despite of fluctuating temperatures during the day- and night time, the large mounds are not just heaps of dirt but in fact incorporate elaborate ventilation holes and air ducts and air pockets, which drive natural ventilation through convection. And remember, these structures are build by a million tiny insects that behave in an organized manner to come up with an architectural masterpiece, every time.
So no wonder that Mick Pearce, born and raised in Southern Africa, was inspired by the the termite mound to design a building in tropical Harare that would have similar features.
Up until recently two models for the termite mound function were proposed and commonly accepted. In case of mounds that were capped a “thermosiphon flow” was created – basically hot air created by the nest rises to the top of the mound where it gets refreshed and is supplied with water vapor through the porous mound walls. This denser air then is forced down below the nest, where the cycle is repeated.
Thermosiphon flow model in which ventilation is driven by heat. (Drawing by Marianne Alleyne, based on Turner & Soar, 2008)
The second model applies to mounds that have a chimney at the top of the mound that opens to the outside. This arrangement creates induced flow, also called the stack effect. The chimney breaks the surface boundary layer and is exposed to higher wind speeds compared to inlets on the ground. The unidirectional flow draws fresh air from near the ground into the nest, where it passes on through the chimney and ultimately to the outside.
Induced flow model in which ventilation is driven by wind. (Drawing by Marianne Alleyne, based on Turner & Soar, 2008)
The architects who designed the Eastgate Centre building tried to incorporate both the thermosiphon and induced flow principle into their design. The building has an extensive tube system within the walls and floors that move air trough the building. Heat generated within the building, along with stored heat within the structure, creates a thermosiphon-effect that draws air up and through the rooftops where large chimney stacks are located. These tall stacks are essential for creating an induced flow.
Detail of tube system within Eastgate Centre building’s walls and floors (Original picture at http://harare.usembassy.gov).
When popular stories about the Eastgate Centre building say that the building works on the termite mound principle they ignore the fact that the building uses low capacity fans during the day, and high capacity fans during the night to keep the air from being too stagnant, effectively replacing the hot air that builds up during the day with cool air during the night. This works well, and avoids having to use expensive air-conditioning technology, but needless to say, no termite mound utilizes fans.
The use of fans within the Eastgate Centre. Smaller fans run during the day-time hours (left). They keep the environment within the building comfortable while the walls store the heat from the outside. Larger fans run during the night-time (right), these fans pull the stored heat out of the walls and push the heat out through the ducting in the ceiling and walls. By the next morning the walls are ready to again store heat (Drawing by Marianne Alleyne).
Turner and Soar actually measured temperatures in and outside of the mound. It turns out that while termites may be able to dampen temperatures during the day the nest itself actually closely tracks the soil temperature, which ranges from 15 degrees C in the winter and 31 degrees C in the summer. Mounds have clearly a large thermal capacity, but their architecture, their infrastructure, and their ventilation have little to do with the internal temperature at any given time.
Nest temparature (blue) and ground temperature at 1m depth near nest (red) Drawing by M. Alleyne based on Tune r& Soar, 2008))
In addition, whereas induced flow might work well in tall buildings because the likelihood of a boundary layer gradient between locations is pretty high, it has been shown that induced flow rarely operates in termite mounds, even open-chimney mounds, since they are commonly only about 6 ft tall. There is also no evidence that mound ventilation and nest ventilation are indeed linked. How respiratory gases are moved from the nest to the mound, and fresh air from the outside, through the mound, to the nest, is not well understood.
Now the Eastgate Centre, and other large buildings since then, accomplished what they set out to do – they saved in construction costs (HVAC systems are very, very expensive) and they save on operating costs, all while keeping the inhabitants comfortable. It probably does not matter that they were based on incorrect science, but it does matter that the misconception gets repeated over and over again. If biomimicry and/or bioinspiration want to be considered legitimate fields of study, and not just a feel-good endeavor, then the science that the field is based on has to be solid.
The termite mound should still inspire developers and architects because at it turns out, if we view the termite mound as the analogue of our own, respiratory system (lungs) then we still should be able to design “breathing” buildings that have that walls serve more as membranes rather than barriers.
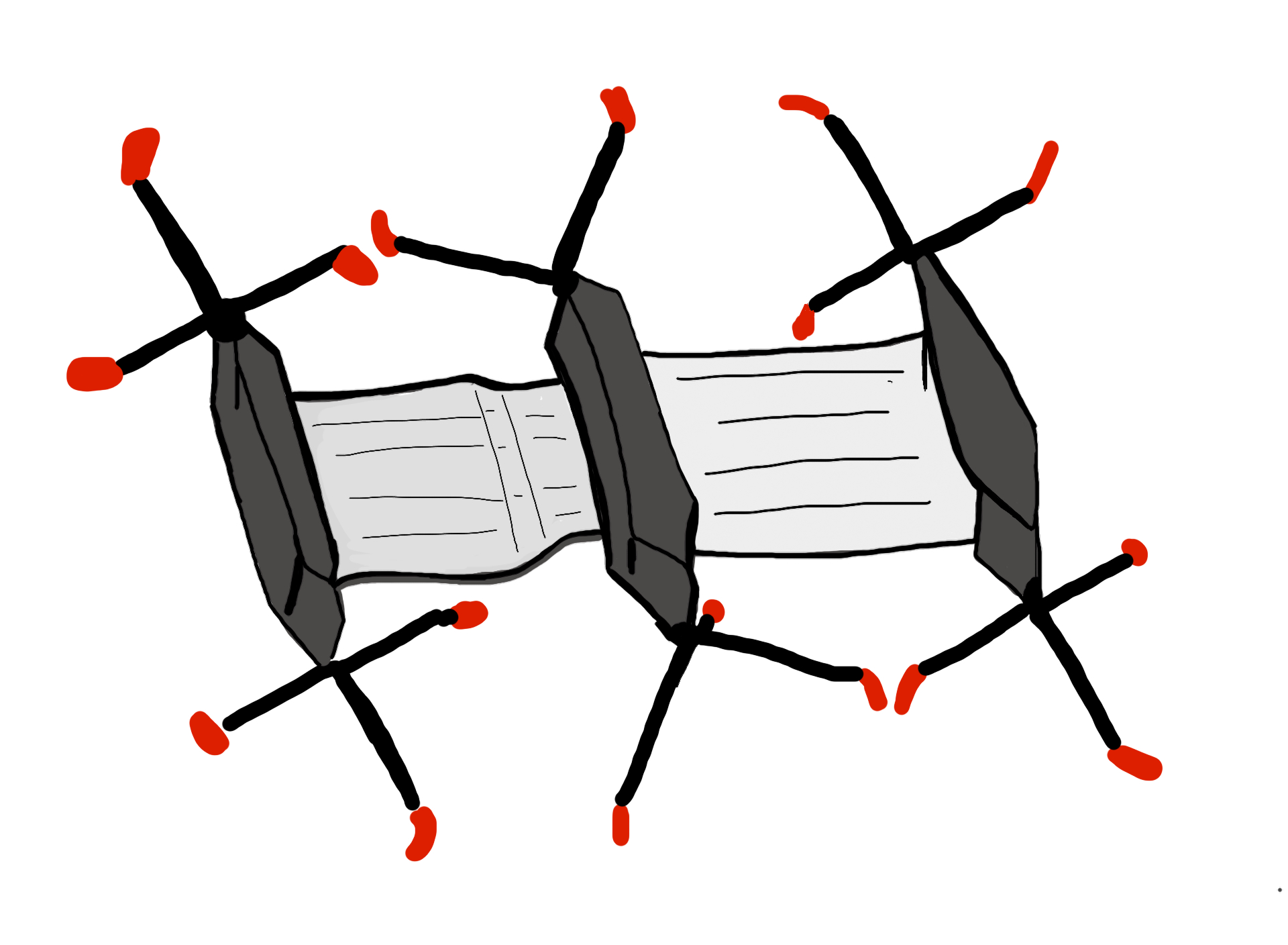
Turner and Soar not only took relatively simple measurements of the temperatures, they also made plaster casts of the tunnel network of M. michaelseni mounds. They then created horizontal slices of the plaster filled mounds for easier recreation of future 3D models.
Based on these models, which showed actually very little continuous mixing between the air coming down from the above-ground structure and the air from the underground nest, they propose that the termite mound of M. michaelseni is in fact a functional analogue of a lung. And much like the lung the termite mound is far more complex than the simple models we have been using (for an excellent explanation comparing functional organization of both lungs and termite mounds please refer to Turner and Soar, 2008).
Comparison of the functional organization of mammalian lungs and the termite mound. There are areas of forced convection (large tidal flows) (in red), areas where smaller tidal flows dominate creating a mix of diffusion-convection (in blue), and then the areas where diffusion dominates (in green). (Drawing by Marianne Alleyne, based on Turner & Soar, 2008)
This complexity opens up new avenues for bioinspired design based on the termite mound. Termites erect walls that are actually interface directly with the outside and indoor environments – the walls themselves let gases and energy through, it does not form an impenetrable barrier. Buildings that are designed based on correct termite mound architecture should incorporate porous walls and cladding (~skin) systems that incorporate tubes/tunnels through which air and energy can flow. Maybe we can design living buildings that are part of our extended physiology, as much as the termite mound is part of the living-system called the termite colony.
Resources:
I cannot recommend the Turner and Soar paper from 2008 enough. It is a wonderful read. The authors touch on more topics than I mention here (homeostasis, for instance). And again, their debunking of previously held beliefs is done in a way that should be emulated.
J. Scott Turner and Rupert C. Soar. 2008. Beyond biomimicry. What termites can tell us about realizing the living building.
The same authors also wrote a book chapter on a similar topic. But I have not yet been able to locate: Beyond biomimicry. What termites can tell us about realizing the living building. Chapter 15 in: Industrialised, Integrated, Intelligent Sustainable Construction. ISBN 978-0-86022-698-7. Ian Wallis, Lesya Bilan, Mike Smith & Abdul Samad Kazi (eds). I3CON/BSRIA. London. pp 233-248.
J. Scott Turner has also converted this work into very informative video lectures, such as:
Learning from nature: Termite mound lungs and the implications for breathing mines.
For great footage of how the termite mound models were created check out these videos.
I am not, by a long shot, the first to point out how the termite mound is NOT (yet) the perfect poster-child for bioinspiration and biomimicry. Here are two other blog-posts that discuss this very topic:
Miscellaneous – things I learned about social media while writing the blog-post.
Silkworm pavilion
I ended last month’s “Bits and Bytes” with a mention of a project that was making the rounds under the heading #biomimicry. I am still not sure if this really falls under biomimicry or bioinspiration, but during the month of June it kept popping up on Twitter, probably because it is just very, very cool. The project received extra attention because the project was on display in Boston, the city where the Biomimicry3.8 Education Summit and Global Conference was held during the month of June – so cross-pollination for all!
The Mediated Matter group, under the guidance of Professor Neri Oxman, studies additive fabrication techniques (such as 3D printing) and tries to scale some of them up to, for instance, building-size structures. One of the projects involves mobile swarm building where small robots could potentially build large structures. For this project, which is ongoing, they are currently studying how silkworms (the caterpillars of the moth Bombyx mori) can inform this type of building technique.
Silk Pavilion information can be found on the Mediated Matter website here (tools: swarm printing), and here (environments: silk pavilion), and here (news: silk pavilion).
Last month’s Bits and Bytes started off with the perfect insect-inspired story: an engineered compound eye from a University of Illinois lab. Great story to lead off the inaugural issue of a recurring blog feature – considering I cover insects, engineering and work at UIUC. Turns out that some of my ommatidia neglected to notice another story very similar to the one I covered. Last month the Laboratory of Intelligent Systems at EPF-Lausanne also created a miniature curved artificial compound eye.
Size comparison between CURVACE (Curved Artificial Compound Eye) and a dragonfly. Image: courtesy Dario Floreano / Swiss Federal Institute of Technology. http://curvace.org/
Maybe the manufacturing technique and the material used are not as cutting-edge as those used in the Rogers’ lab, but it seems to me that the final creation is a lot more like an insect compound eye.
The design consists of three planar layers of separately produced arrays, namely, a microlens array, a neuromorphic photodetector array, and a flexible printed circuit board that are stacked, cut, and curved to produce a mechanically flexible imager.
Reference: Floreano, D., R. Pericet-Camara, S. Viollet, F. Ruffier, A. Brückner, R. Leitel, W. Buss, M. Menouni, F. Expert, R. Juston, M.K. Dobrzynski, G. L’Eplattenier, F. Recktenwald, H.A. Mallot & N. Franceschini. (2013) Miniature curved artificial compound eyes. PNAS V110 (230, pp 9267-9272. DOI: 10.1073/pnas.1219068110
Robotic insects were again quite popular this month, or at least on Twitter they were. As I covered in an earlier post (The Dawn of the Artificial Coprophages) engineers have been interested in building robots that move and behave similar to insects for quite a while. It first started with terrestrial locomotion, but now we also see insect-inspired robots that can swim, walk on water, dig, jump and (The Holy Grail) fly.
Here are a few insect-inspired robots that came across my computer screen this week:
Lentink reviews how the basic research on aerodynamics of insect flight inspired engineers to build robots using the data obtained by the biologists. And how this cross-pollination occurs at the same time as micro-manufacturing techniques are being developed. The article culminates with a recent publication from Rob Wood’s Harvard Microrobotics Laboratory.
Ma, K. Y., Chirarattananon, P., Fuller, S. B. & Wood, R. J. 2013. Controlled flight of a biologically inspired, insect-scale robot. Science. http://dx.doi.org/10.1126/science.1231806
This article was also covered by Peter Forbes (author of the Gecko’s Foot) on his blog.
Monosson does not mention insect or insect-inspired robots specifically but she does wonder if life/AI can evolve from wires and plastic. Considering all the work that is being done on insect-inspired robots I assume that we will soon find out.
Another story that received a lot of twitter-buzz this month was the one about Morpho butterflies serving as inspiration for anti-counterfeit money. The fact that Morphos (and many other butterflies and moths) use structural colors has been known for some time. Engineers and Material Scientists have also been interested, for quite a while now, in manufacturing materials that incorporate nanostructures similar to those on the butterfly wing. So that begs the question, why is this big news now? Probably because of a strategically placed corporate press release. It is interesting to see that news can spread quickly via social media (see silkworm pavilion story, for instance) but also that the story never really goes away.
Another example of how not-really-news becomes (again) a relatively big story is the article by Rafe Sagerin on how biomimicry can help us stay ahead of hackers. This article was basically a synopsis of his 2012 book called “Learning from the Octopus” which was fun book to read – but did not feature near enough insect examples. This article mentions only one insect (the stingless bee, go figure) but sentences like;
“The best bet is to do what the most successful organisms on Earth do — accept the risk and adapt to the changes”
immediately makes me think of insects since they are so very very successful and ubiquitous.
That this particular article became so popular this month showed me that getting your core-message (look to nature to fight terrorists, diseases, hackers) into a magazine where the topic is not typically discussed (Harvard Business Review) will help reach a lot of new sets of eyes and will familiarize more people with the terms biomimicry and bioinspiration.
Miscellaneous
Lessons I learned from Twitter and this blog-post during the month of June:
There is a lot of cool stuff out there about insect-inspired technology. I need to become better about cataloging all the awesomeness as it becomes available during the month. (Any tips?)
There is so much cool stuff out there that sometimes I miss something, especially if the topics are similar to each other. Last month I assumed the engineered compound eye stories were one story, but it turns out that they are actually two different approaches.
Some stories seem like new but they may actually be receiving a “second wind”, because a journalist/science writer revisits the story or because of a simple retweet. Some stories therefore never really disappear.
Placing a story in a non-traditional media outlet will result in a whole bunch of new eyes seeing the work, and that is a good thing.
And after all these years I am still unsure if certain areas of research really fall under bioinspiration or biomimicry (for example, the silkworm story or biofuels). And that is OK. Not everything will fit neatly in a box, even though the scientist-side of me wishes it would.
The ability to manufacture devices at the very small, or micro, scale brings with it interesting applications possibilities and surprising challenges. For instance, certain physical phenomena may not be generally encountered with doing macro scale experiments on the lab bench but are dominant when experiments are done with a micro scale “lab-on-a-chip.” The field of microfluidics deals with the behavior and manipulation of fluids, both liquids and gases, that are constraint to a small scale (1 to several hundred micrometer). The physical properties of fluids at this scale are particularly interesting and could possibly be exploited. For instance, fluids at a certain scale are no longer practically compressible. Also, it is difficult to maintain pressure or pressure-driven flow within a micro scale fluidic system. One example is the phenomenon of back-flow whereby fluid moves in one direction when pressure is applied only to have it reverse and flow back when the pressure is released. Due to the physics of scaling laws, these various non-intuitive phenomena are less severe when transporting gasses than liquids in a micro scale device. For instance, gravity has no influence on gasses and diffusion of gasses at the micro scale is VERY fast. Therefore, gas species can mix through diffusion with simple laminar flows at low Reynolds numbers.
In nature microscale liquid and gas flow is quite common. Maybe engineers should look at insects to see how fluids can most effectively be moved around at the microscale. The insect’s respiratory system, for instance, uses microscale flow transport to achieve an absolutely essential physiological function. The respiratory system at first glance seems so simple – just some gas-filled tubes. At the same time it seems so foreign – no breathing through mouth or nose, no lungs or blood or hemoglobin involved (at least not at first glance). The respiratory system is also elegant in its simplicity through ingenious feedback loops – fine-tuning through direct nervous innervation or by sensing chemical compositions of the blood (called hemolymph in insects).
This system that is key to maintaining homeostasis is also as varied as insects are varied. Ingenious adaptations are seen in insects that live in different habitats – within soil, in water, in arid climates, in cold climates, within a host, while pupating, while flying, while walking, etc.
In this post I’ll focus mostly on the “generalized” terrestrial insect.
The insect’s tracheal system
The insect’s respiratory system’s main functions are to carry oxygen and carbon dioxide to and from the tissues. The gas-filled tubes, called tracheae, start at the spiracles, which are paired openings in the cuticle that can occur in almost every or only a few segments (depending on the insect species). The spiracles are innervated by the nervous system and can be open and closed through muscular action.
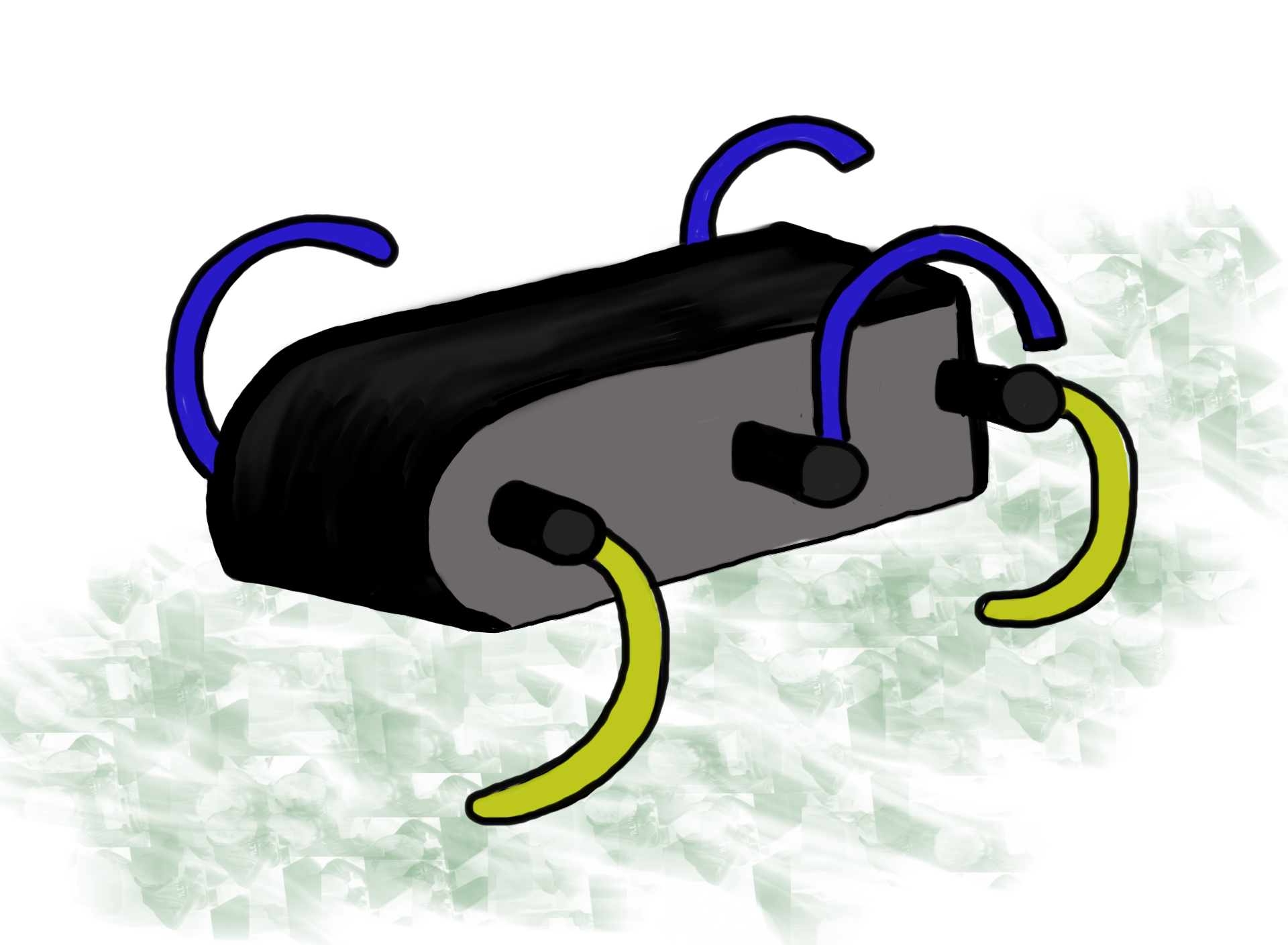
Spiracles of a lepidopteran larva. Two are visible in this picture; they appear as black openings surrounded by yellow ring. (Picture by Geoff Gallice)
Inside the insect the tracheal systems starts out as two or more longitudinal trunks, which further into the insect branch into smaller and smaller channels. Tracheae range in diameter from a few mm to 1 μm. The smallest branches are called tracheoles, which range from 1 to 0.1 μm in diameter. The tracheoles’ tips are in contact with cells and are often fluid-filled. This means that at the tips of the tracheoles oxygen has to cross first a liquid, then the tracheolar walls, then cross the plasma membrane of the cell and finally move through the cytoplasm of the cells to reach the mitochondria. Carbon dioxide makes the same trek, but in the opposite direction.
Tracheal System (Drawing by Marianne Alleyne)
Cross-section of the tracheal system of “general” insect. Other insects may have more than 2 longitudinal trunks and/or air sacs and/or no spiracle functional in a particular segment. (Drawing by Marianne Alleyne)
Cross-section of tracheal tube. Shown is an epidermal outer layer bathing in hemolymph, an inner intima layer, and taenidia, which reinforce the intima layer (much like a car’s radiator hose is reinforced by coil of steel). (Drawing by Marianne Alleyne)
In order to get oxygen to move into the cells and carbon dioxide to move out of the body, all while limiting energy use, insects rely on diffusion and convection.
Diffusion: passive movement of molecules down their concentration gradient.
Oxygen diffusion rate is more rapid in air than in liquid, and because the tracheal system consists mostly of air-filled tubes the gas-exchange by diffusion is extremely rapid and quite substantial. While the cells use the oxygen, the partial pressure of oxygen decreases near the tips of the tracheoles, which then draws in more oxygen from the tracheae and the air outside of the insect. Diffusion is of critical importance at the tips of the tracheoles, but it can only work at micrometer distances since at this point the oxygen has to move through a liquid. This is why almost every cell in the insect is at close proximity to a tracheole tip. The cells that require high respiratory rates (flight muscle cells, for instance) may even have multiple tracheole tips invaginating the cell wall.
The prominent Danish physiologist August Krogh devised an early model (1920) of grasshopper respiration that was based on oxygen partial pressures and suggested that simple diffusion could provide adequate gas exchange in insects. This claim had a huge impact on the field early and quickly became dogma, despite some contemporary contrary evidence. In 1964 “a next-generation Dane” Torkel Weis-Fogh showed that in larger or more active insects the distance between the spiracles and the tips of the tracheoles was too great and that diffusion was insufficient if the insect was active. So what other mechanisms do insects use to move oxygen and carbon dioxide through the tracheae?
Convection: bulk movement of a gas or a liquid driven by differential pressures.
Compared to diffusion, convection can achieve greater gas exchange rates over longer distances. The trade-off is that it requires more work and thus more energy.
One type of convective air-movement seen in insects is, confusingly, termed suction ventilation. This movement of air is primarily achieved by opening and closing, or shielding differentially, a subset or all of spiracles at different parts of the body. This creates a pressure difference between the spiracles and the tracheal ends and thus creates passive airflow.
Autoventilation is achieved by using wings or legs to pump air through the body.
Abdominal pumping, using the muscles in the abdomen to pump air.
Another, more important, type of convective air movement is achieved by the tracheae (tracheal compression), and their associated air sacs (tracheal sections that are not reinforced by taenidia), collapsing and inflating.
Digital image of silkworm Bombyx mori trachea with taenidea (100X mag) by Paul Joseph, Birmingham, UK.
With novel imaging and measurement techniques researchers have shown that rhythmic tracheal compression is quite common. Synchrotron x-ray imaging has been used since 2003 to visualize tracheal structures in living insects. We can now see that muscles compress the exoskeleton of the insect, which results in air moving through the tracheae since the tracheae and air sacs are collapsing. These studies, which are now linked with real-time flow-through respirometry, show that rapid cycles of collapse and re-inflation in a variety of insects’ tracheal systems – especially in the head and thorax regions – results in large changes in tracheal volume and are major components of ventilation in insects. Even the different ways the tracheal system can compress and re-inflate is probably as varied as insects themselves.
Using the insect’s tracheal system as inspiration for microscale flow devices
The impetus for this blog post was a recent paper I came across entitled: Selective pumping in a network: insect-style microscale flow transport, by Yasser Aboelkassem and Anne E. Staples. The topic of microscale flow transport is of interest to engineers because there are many applications that need precise flow control of just microliters of gas or fluid. Microscale flow devices or microfluidic devices are already around us and are assumed to change research tools in the life sciences, aid drug discovery and chemical analyses, and revolutionize health diagnostics. Modern DNA sequencing systems (Fluidigm, Illumina, Roche (454), etc.) rely on microfluidics. “Droplet” analyses also rely on microfluidic technologies. Cheap paper-based microfluidic immunoassay tests will become important for healthcare in the developing world (Diagnostics for All). And those are just a few of the applications.
Insect tracheal systems have been of particular interest to engineers now that we are able to visualize this physiological process (moving fluid and air through tubes) in action. Insects are able to move fluid and air with precision with minimal energy requirements and little input from outside of the body. All characteristics that would work well within a “lab-on-a-chip”.
Insects use contractions at very specific places along the tracheal system, which probably affords them more precise control than if just relying on the more crude pressure difference approach. Through these small, but targeted, compressions the airflow to specific areas of the body can be achieved. This is apparently not an easy task to achieve in small-scale devices, and one can appreciate that by imagining larger tubes.
Imagine a garden hose…(Image by zen Sutherland)
Imagine a garden hose. By stepping onto the hose at one point the fluid or air inside the hose will be displaced to either side. As soon as you step off the hose the fluid or air will rush back (=back-flow). How can one create directional flow of a particular speed or pressure? By stepping at multiple spots along the hose, sometimes closing the hose off completely and other times only partially. Stepping off of the hose according to a certain sequence will also result in a directional flow. Of course, this is just one hose. Now imagine doing this with hoses that are branching into thinner hoses, that have multiple openings, with many more feet that can step on and off of them…and then think of a miniaturized version of that and you will see the insect’s tracheal system.
This is what Aboelkassem and Staples tried to model in their paper. The fluid dynamics research group headed by Staples at Virginia Tech’s Department of Engineering Science and Mechanics is apparently closely aligned with the biomechanics research lab of Jake Socha at the same departments. The Socha lab is one of the labs that has used synchrotron x-ray imaging to study insect respiration. The theoretical and computational model proposed by Aboelkassem and Staples incorporates rhythmic wall contractions along a network of tubes (in this case an 8-armed network) without the need for valves. The model’s tubes/channels are squeezed at different places along the tube. The contractions were actuated to move with different time lags from each other. In this way the researchers showed that fluids can be transported and flow velocity, pressure and direction can be controlled.
Simplified insect tracheal network modeled by Aboelkassem & Staples. The “tracheae” can be collapsed at multiple points within the network (red arrows). The collapse motion protocol can be varied by including time phase lags. The result is precise movement of the liquid or gas (green arrows).
We have learned from insects that localized rhythmic tracheal contraction is an effective way to move gas through micro-tubes. Aboelkassem and Staples claim to have been inspired by the insect’s tracheal system to model “selective pumping in a network”. They show, by using theoretical (low Reynolds number) flow analyses and computation fluid dynamics, that the network might also enable fluids to be transported precisely into particular tubes without the use of mechanical valves – at least through an 8-armed tracheal system. It will be interesting to see if this research group will continue to follow the insects’ lead by adding more branches/channels, by adding air sacs (which seem to be very important in insects), by making the model 3D, by adding actively opening and closing ports/spiracles to the outside, and by incorporating the diversity of insect respiratory systems into different models. If done correctly, and by using microfabrication techniques that can make flexible micro- and nano-scale tubes, it is likely insects can make a contribution to the rapidly growing technological field of microfluidics.
Resources and References:
For a more detailed description of the insect’s respiratory system:
Harrison, J. F., J. S. Waters, A. J. Cease, J. M. VandenBrooks, V. Callier, C. J. Klok, K. Shaffer and J. J. Socha. (2013) How Locusts Breathe. Physiology V28:(1) 18-27. DOI:10.1152/physiol.00043.2012
Harrison, J.F. (2009) Respiratory System. Pages 889-895 in: “Encyclopedia of Insects, 2nd ed.”. Editors, V.H. Resh and R. Carde, Elsevier, San Diego.
Harrison, J.F. (2009) Tracheal System. Pages 1011-1015 in: “Encyclopedia of Insects, 2nd ed.”. Editors, V.H. Resh and R. Carde, Elsevier, San Diego.
For description of how synchrotron x-ray imaging is used to study insect respiration:
Westneat, M. W., O. Betz, R. W. Blob, K. Fezzaa, W. J. Cooper and W. -K. Lee. (2003) Tracheal respiration in insects visualized with synchrotron x-ray imaging. Science V299, 5606, pp. 558-560. DOI: 10.1126/science.1078008
Westneat, M.W., J.J. Socha and W.-K. Lee. (2008) Advances in biological structure, function and physiology using synchrotron x-ray imaging. Annual Review of Physiology V70: 119-142. DOI: 10.1146/annurev.physiol.70.113006.100434
Socha, J.J., W.-K. Lee, J.F. Harrison, J.S. Waters*, Fezzaa, K. and M.W. Westneat. (2008) Correlated patterns of tracheal compression and convective gas exchange in a carabid beetle. Journal of Experimental Biology V211: 3409-3420. DOI: 10.1242/jeb.019877
Socha, J.J., T. Förster and K.J. Greenlee. (2010) Issues of convection in insect respiration: Insights from synchrotron x-ray imaging and beyond. Respiratory Physiology and Neurobiology V173S S65–S73. DOI: 10.1016/j.resp.2010.03.013
Waters J.S.,W.-K. Lee, M.W. Westneat and J. J. Socha. (2013) Dynamics of tracheal compression in the horned passalus beetle. American Journal of Physiology:Regulatory, Integrative and Comparative Physiology V304(8) R621-7. DOI: 10.1152/ajpregu.00500.2012
Aboelkassem, Y. and A. E. Staples. (2013) Selective pumping in a network: insect-style microscale flow transport. Bioinspiration & Biomimetics V8(2) 026004. DOI: 10.1088/1748-3182/8/2/02600
Special thanks to Dr. Paul Kenis for taking the time to discuss microscale flow transport with me. I appreciate his patience with me – clearly a novice. Bedankt!
Some of you might be thinking that the topic of this blog is rather narrow: “How can she possibly sustain this blog because she is going to run out of topics at some point…soon”.
Or is that just the voice in my own head talking?
I only have to remind myself that I have chosen a taxon, the Insecta, which is extremely diverse and that new species are being discovered and described every day, so it is very likely that I am going to be occupied for a while. Insects have adapted to many different environments, often through very novel and varied (compared to mammals) adaptations. Inspiration for innovation is bound to be found in common insects, but also in the obscure. And if I adopt the often ignored non-insect arthropods such as ticks, mites, spiders, etc., then I will be set until retirement. At the same time imaging and manufacturing techniques are making the small visible and producible so advances in engineering are helping me stay off the streets too.
In this inaugural “Insect Bits & Bytes” post I highlight some of the research I came across on Twitter (#biomimicry or #bioinspiration) during the month of May. These studies all involve new technologies that were inspired by insects or basic discoveries about insect biology that could lead to new innovations.
I have compiled a list of links to this work, as well as to coverage of the research by some of my favorite science writers. I hope to do the same at the end of every month.
Compound eye-inspired camera
The biggest insect-inspired technology story came from the University of Illinois at Urbana-Champaign (my home institution). John Roger’s material science lab was inspired by the insect eye to develop a new digital camera.
New digital cameras exploit large arrays of tiny focusing lenses and miniaturized detectors in hemispherical layouts, just like eyes found in arthropods. Photo credit: John A. Rogers, UIUC
These hemispherical cameras depend on the manufacturing technique that has been perfected in the Roger’s lab – manufacturing flexible electronics.
Engineers have tried to manufacture compound eyes before. In 2006 UC Berkeley’s Luke Lee fabricated an artificial compound eye in his lab. He created thousands of closely packed light-guiding channels leading to pin-head-sized polymer resin domes and then topping each dome with its own lens. Each individual unit is very similar to an insect’s ommatidium (the individual unit of the compound eye). The fabrication method itself was based on the developmental stages of the insect, and resulted in a 3D artificial compound eye that is similar in size, shape and structure to the insect’s compound eye.
In my opinion, because of how the “eye” is manufactured and functions, Lee’s artificial eye is closer to its model than this new bioinspired eye from the Roger’s lab. Time will tell if, by adding engineering shortcuts in manufacturing, and by using materials that work better with how we currently use electronics, a more useful camera or sensor is created.
Reference: Song, Xie, Malyarchuk, Xiao, Jung, Choi, Liu, Park, Lu, Kim, Crozier, Huang & Rogers. 2013. Digital cameras with designs inspired by the arthropod eye. Nature http://dx.doi.org/10.1038/nature12083
Over the years it has been exciting to see how small engineers can make flying robots. Research by people like Michael Dickinson on insect aerodynamics have helped engineers such as Ron Fearing and Rob Wood to develop microrobots that can fly. This past month we learned that the Wood lab at Harvard’s Wyss Institute has now manufactured a controllable robot, the size of an insect, that can fly. (Note: the manufacturing process for these types of robots is really cool too.)
One of the major remaining challenges, before microrobots will be used on a grand scale, is to get them enough power to walk, run, swim and/or fly for an extended time (note the tether in all the flying minirobot pictures and videos). There is just not enough room on a small robot to incorporate conventional batteries, or even smaller lightweight battery sources like a coins cell or solar panels. Future advances may involve biological motors as power sources. Maybe we can even learn more about basic insect flight energetics (a very interesting topic) and incorporate what we learn about basic insect physiology into microrobots.
Reference: Ma, K. Y., Chirarattananon, P., Fuller, S. B. & Wood, R. J. 2013. Controlled flight of a biologically inspired, insect-scale robot. Science. http://dx.doi.org/10.1126/science.1231806
Cicadas all over the news these days. The East Coast of the US is in the midst of the 17-year periodical cicada emergence. This year cicadas are apparently also of great interest to those studying biological materials at the nanoscale. Earlier this year it was reported that nanopillars on clanger cicada wings can tear bacterial membranes apart. One can think of interesting applications for engineered materials that incorporate similar structures.
This month another study showed that cicada wings are also extremely hydrophobic; droplets pretty much jump off of the surface. The wings are thus self-cleaning. Again, one can think of multiple applications for an engineered hydrophobic material based on the cicada wing. Then again, there are many other examples of biological materials that have similar characteristics: lotus leaf, Namib beetle, etc. One interesting idea that Charles Choi brought up in his article (link below) is the use of cicada-wing technology in power plants. Jumping droplets would help dissipate heat.
Reference: Wisdom, K. M., J. A. Watson, X. Qu, F. Liu, G. S. Watson & C-H. Chen. 2013. Self-cleaning of superhydrophobic surfaces by self-propelled jumping condensate. Proceedings of the National Academy of Sciences of the USA. http://dx.doi.org/10.1073/pnas.1210770110
The coverage of this story was, and still is, plagued by a #TaxonomyFail (pointed out to me by @BrianTCutting). Most of the coverage associated with this story showed a picture of a wet fly, sometimes a wet fly that was upside down. Soon I hope to add a picture to this post of a wet Brood II cicada. Stay tuned.
Thin film interface inspired by moth eyes
The insect eyes have it, again.
Moth eyes were the inspiration for a new multilayered material which may find application in optoelectronic devices such as solar cells. For at least 40 years we have known how nature solves the problem of light reflection. We only now have the imaging and manufacturing capabilities that will enable us to engineer and produce materials that mimic the most effective nanostructures.
Moths are generally nocturnal and any light the eye can “harvest” is a plus, reflection of light needs to be minimized.
Moth eyes reflect very little incident light. (Image by Daniel Meyer)
The “moth eye” principle was first described in 1973 by Clapham and Hutley. Their electronmicrographs showed that the surface of corneal lenses of moths are covered with conical nanostructures and it was proposed that these structures suppressed interference (reflection).
Over the past decade nanostructured materials mimicking the moth eye have been manufactured through techniques such as ion-beam etching, but application was limited because the material could only be manufactured at a small scale. Recently researchers at North Carolina State University were able to manufacture interfacial nanostructures protruding from a silicon layer into a overlaying thin film and thus eliminated interference effects. It remains to be seen if the manufacturing technique proposed in this recent work can be scaled up to produce consistent nanostructures at a reasonable cost.
Reference: Yang, Q., X. A. Zhang, A. Bagal, W. Guo & C-H Chang. 2013. Antireflection effects at nanostructured material interfaces and the suppression of thin-film interference. Nanotechnology http://dx.doi.org/10.1088/0957-4484/24/23/235202
Physicists and biologists worked together to explain how fire ants tunnel through the ground. The types of descriptions of locomotion will help engineers build more useful robots.
Obviously legs are important for locomotion on land, however, functional feet may not be just the distal end of a leg (cockroach). Also, appendages such as tails, are essential for dynamically stable locomotion (gecko). These types of biomechanical principles have already been incorporated into robots. Now a recent study from Georgia Tech shows that additional appendages, antennae, do not just serve as chemical or mechanical sensors. When falling the antennae help the ant grab onto the tunnel wall. Civil engineers might also learn from this biological example since ants build tunnels close in diameter to their own body length, no matter what the substrate, so that all legs and antennae can help get a grip when falling.
Reference: Gravish, N., D. Monaenkova, M. A. D. Goodisman & D. I. Goldman. 2013. Climbing, falling and jamming during ant locomotion in confined environments. Proceedings of the National Academy of Sciences. http://dx.doi.org/10.1073/pnas.1302428110
Turns out that the animal with the best hearing is the greater wax moth (one of the many scourges of bee keepers). Moths have a tympanum on either side of the abdomen. Each tympanum is innervated by just two sensory receptors. These receptors start firing at the slightest displacement of the “ear drum”. Turns out that the greater wax moth can sense displacement caused by frequencies up to 300 kHz. In addition, this type of auditory system works at a wide range: from 20 kHz up to 300 kHz. Engineers are keen on building a mechanoreceptor as sensitive to ultrasound as this, and with materials and structure as “basic” as a moth’s ear.
Interestingly ultrasonic sensors are preferred over photoelectric sensors in certain situations – now bioinspired technologies based on the moth eye (see above) and the moth ear may blur those distinctions.
An illustration from British Entomology by John Curtis. Lepidoptera: Galleria mellonella
Jumping Robots
Also, everyone’s favorite feisty insect-inspired robot, Rhex, learned to jump.
Reference: Johnson, A. M. & D. E. Koditschek. 2013. Toward a vocabulary of legged leaping. Proceedings of the 2013 IEEE Intl. Conference on Robotics and Automation. http://kodlab.seas.upenn.edu/Aaron/ICRA2013
A history of insect-inspired walking robots and how they “evolved”.
Earlier this year our Department of Entomology at the University of Illinois at Urbana-Champaign hosted the 3oth Annual Insect Fear Film Festival. This year’s theme was InsX-files and combined two “alternative” communities – those passionate about insects and those passionate about a TV show that has not been on the air for years, the X-files. You might be surprised to learn that even in the Midwest these communities can fill up a big lecture hall no problem (I was).
30th Insect Fear Film Festival Promotional Design (Theme: Ins-X Files). Designed by Joseph Wong for Illinois’ EGSA.
Special guests at the festival were series creator, writer, producer and director Chris Carter and the writer of some of the most popular episodes of the show Darin Morgan. (For scenes from the festival check out the tweets at the end of this post)
One of those episodes, entitled: “War of the Coprophages” (1996), was shown at the festival. This particular episode has achieved cult status here at Illinois because it features the character Bambi Berenbaum, who in her appearance and mannerisms is exactly like nothing like our fierce leader and Department Head May Berenbaum. (Morgan consulted various books authored by May and thus decided to give the fictional USDA scientist a name that honored her).
Another reason why I love this episode (besides the fact that it is basically a fantastic piece of suspenseful science fiction writing) is that it featured insect-inspired robots. In short, a town somewhere gets overrun with cockroaches that may be killing people. Some of the roaches appear to be mechanical rather than biological. Agent Mulder decides to visit a researcher named Ivanov at the Massachusetts Institute of Robotics to get a better idea about what engineers are up to.
(Mulder walks down the stairs and then a hallway. A small, insect-shaped robot walks down the adjacent hallway (and manages to erase the title of the location as he does somehow.) Mulder watches as it walks into a room, then comes out and looks at him. As Mulder takes a few steps towards him, the insect back away. Mulder follows the insect to a laboratory where the insect robot disappears. Mulder hears a similar whirring and turns around to see Doctor Ivanov approaching him on his wheelchair. The scientist talks out of a microphone that is near his throat. A laptop computer is hooked onto the wheelchair.)
IVANOV:For decades, my colleagues in artificial intelligence have attempted to create an autonomous robot. By struggling to give their machines a human-like brain, they have failed. A human brain is too complex, too computational. It thinks too much. But insects merely react. I used insects as my model, not just in design but by giving them the simplest of computer programs. “Go to the object. Go away from the moving object.” Governed only by sensors and reflex responses, they take on the behavior of intelligent, living beings.
MULDER: So this one is just programmed to head towards any object moving within the field of its sensors?
IVANOV: No.
MULDER: Then why is it following me?
IVANOV: He likes you.
MULDER: Your contract is with NASA?
IVANOV: The goal is to transport a fleet of robots to another planet and allow them to navigate the terrain with more intricacy than any space probe has done before. It, it sounds slightly fantastic, but the only obstacle I can foresee is devising a renewable energy source. In any case, this is the future of space exploration. It does not include living entities.
Screenshots from the X-files episode “War of the Coprophages” showing two of the insect-inspired robots featured in the episode.
Genghis
Darin Morgan could have gleaned inspiration from another University of Illinois Entomology Faculty member for ideas about robots, namely Fred Delcomyn who around 1996 was also working on a cockroach-inspired robot. Instead the Ivanov character is clearly based on MIT’s Rodney Brooks, a roboticist who, at the time, wanted to move away from incorporating Artificial Intelligence into robots and instead conceived of robots that were more adaptive to their environment. By programming only simple modules of behavior into the robot, rather than complex reasoning parameters, and let the robot thus react to the environment, Brooks felt that he could build some very functional robots. Much like the ones featured in the X-files episode. Brooks’ mid-nineties robot is named Genghis (~1991). Genghis was the first robot created by the iRobot Corporation (now of Roomba fame) and was intended for possible planetary exploration. Other insect-inspired robots from Brook’s MIT “Insect Lab” includes Boadicea which employs a differential leg design to allow for longer stride frequencies and an increase in speed (the cockroach Blaberus discoidalis served as the model). Both Genghis and Boadicea are robust to failures in leg function, much like animals that can adopt a compensatory gait. Choosing robustness over perfection is a characteristic that shows up again and again in successful bioinspired robot design.
A hexapodal robot named Genghis from the Brook’s lab at MIT (designed around ’91). The robot was also relatively cheap thanks to innovative construction methods. Genghis used 4 microprocessors, 22 sensors, and 12 servo motors. (Picture from NASA.gov)
Around the same time that Genghis became famous other research groups in the United States also started building walking robots that incorporated the biomechanics of actual terrestrially locomoting insects. I review some of the more successful projects here.
Why build robots based on insects?
Insects exhibit behaviors that are considered relatively simple, and thus easier to emulate. The resulting behavi0r may appear complex or purposeful to the observer but it is actually derived from fairly simple rules on how the nervous system perceives environmental inputs and how an internal “neural” pattern generator relays this information to “external” mechanical components (muscle, legs). Of course, “simple” is relative. Different situations may also result in different behaviors (and neural input). For instance, when cockroaches are close to wall they tend to amble, at this speed the animal is very sensitive to nervous feedback from its surroundings, each individual leg is more sensitive to this feedback. When roaches trot really fast across open spaces then a central pattern generator may “suffice”. This CPG generates rhythmic movements in a neural circuit, with little feedback from the environment.
In addition, the exoskeleton and the muscles stabilize insects without the involvement of the nervous system. Hexapod locomotion is dynamically very stable (this claim was conclusively proven by adding jet-packs to roaches…oh, yes, indeedy). Just like its model insect-inspired robots basically uses 2 tripods for locomotion; at any time during the gait the insect has three feet on the ground. The sprawled posture also results in passive stabilization of lateral motion, it is very difficult to push over an insect or a robot using 6 legs. (For a very detailed explanation of these two points please watch this March 2013 seminar by Princeton’s Philip Holmes)
Top figure: Cockroaches walk with a tripod gait: they always keep one tripod of legs (the foreleg and hindleg from one side and the middle leg from the other) in contact with the ground, alternating the tripods as they walk. In the bottom figure are respresented the stance phases of the two different tripods (red or blue) (Drawings by Marianne Alleyne)
Another goal of roboticists has always been to miniaturize their creation. Making cheap little robots is still the goal. Smaller robots can potentially survey areas that are currently not accessible. Also, if you have multiple smaller robots available then you can send more to one area, each carrying cameras and chemical sensors, while a robustness (missing or non-functioning individual robots) is built into the system. As we shall see, miniaturizing brings its own challenges. Insects are small, some insects live in social groups, so there is lots to learn about miniaturization and swarming from them.
It is also much easier to make a robot that has an exoskeleton that is segmented (rather than an animatron that has an endoskeleton). Insects are segmented animals and in some of the insect-inspired robots we see this segmentation too because it increases flexibility.
Since the 1996 episode of the X-files many other insect-inspired walking robots have spawned. Even evolved.
1. RHex – Robotic Hexapod.
Insects such as cockroaches served as the biological inspirations for RHex. Data on bio-mechanics and dynamics of insects maneuvering over rough terrain were obtained by the researchers from the PolyPedal lab at UC Berkeley (Robert J. Full is the primary investigator of the lab) (For full disclosure the author of this blog was once an undergraduate in the PolyPedal lab working on the energetics of locomotion in crustaceans). (Bob also gave 3 very informative TED talks)
Terrestrial animals (bipeds, quadrupeds, hexapeds, octopeds) all rely on a spring-mass system where the limbs (incl. muscle and cuticle) have a spring-like function to help support the animal’s weight over the course of the stride (larger animals have stiffer springs). In addition, the neural control of muscle action during walking and running is linked to muscle stiffness and thus the spring.
The biological data was then used by engineers Dan Koditschek (at the University of Pennsylvania), Al Rizzi (Carnegie Mellon University) and Martin Buehler (then at Boston Dynamics) to build a robust autonomous robot that was able to transverse uneven ground without actual terrain sensing or actively trying to control adaptive maneuvers.
Despite the fact that RHex legs have many degrees of freedom (many legs, joints and actuators), by incorporating real biological data and following the simple rules of a spring-mass system a robot was created that is quick (as measured in body lengths per second), maneuverable, and robust.
One of the most striking advancements of RHex was the compliant legs which were made of materials that helped with dynamic stability, shock absorption, energy efficiency, enhanced gait control, obstacle avoidance, etc. (RHex is now part of the Boston Dynamics robot-thoroughbred stables)
One generalized version of RHex (Robotic Hexapod), the first legged robot to run over uneven terrain, and the first autonomous legged platform to run at speeds above one body length per second. (Drawing by Marianne Alleyne)
Since funding for RHex started in 1998 RHex has evolved into different versions (species?). By 2012 feisty RHex had developed into this:
And very recently it was announced that RHex is also able to leap.
There is also a a cost-efficient education and research version called EduBot.
(For some great pictures and video of RHex click on this Boston Dynamics website).
For an explanation on how you can use biological research (by Joe Spagna and others) done with RHex in your college courses and outreach project click here.
2. Sprawl
Whereas I have always found RHex to have a spunky personality the robot Sprawl to me seemed to have something sinister about it. Must be because of all the wires and (pink!) tubing. Of course, some of the more “evolved” versions have names such as Franken-Sprawl or Sprawlita which does not help them win cuddliest-robot contests.
Member of the Sprawl family. One of the first fully dynamic locomoting hexapods. (Drawing by Marianne Alleyne)
The Sprawl family of robots were created by Mark Cutkosky‘s group at Standford University’s Center for Design Research, again using data from Berkeley’s Polypedal lab. The Sprawl robot incorporates biological principles not only in its leg arrangement and design, but also in its construction and in the material properties of its structure. The robot was made using (then) modern manufacturing techniques (shape deposition manufacturing) to create limbs of the right shape and with the desired material properties, like stiffness at certain critical points. Early Sprawl robots used pneumatic actuators, whereas the later iSprawl robots used electric motors and flexible cable drives. The final result is a sturdy and super-fast robot that resembles a scurrying cockroach. Over the years Sprawl (now called Sprawlettes) have become smaller and smaller (currently you can hold one in the palm of your hand) and they can now be batch-manufactured.
3. Whegs
Another successful collaboration between biologists and robotocists can be found at Case Western Reserve University. Since the 1990s Mechanical Engineer Roger Quinn’s group has used data from neuroscientists such as Roy Ritzman to build cockroach-inspired robots that can walk and climb (for instance, the hexapod Robot III from ~1999). One line of robots is the WHEGs family of robots which use a Wheel-Leg hybrid. The robot was inspired by the European Space Agency’s Prolero robot and RHex, but instead of using 6 motors to drive individual legs (as RHex has) it only uses one powerful one, which can distribute its power to all or just a few of the legs.
Representation of an early model Whegs: this robot comines the advantages of wheels (speed) and legs (maneuver over obstacles) (Drawing by Marianne Alleyne)
The later models of Whegs mimic cockroach maneuverability to manage uneven surfaces. In addition, cockroach behavior during locomotion is copied by adding a variety of sensors. Cockroach rely on antennae to guide them over and under obstacles. Whegs robots are fitted with mechanical antennae that mimic the movements of the cockroach antennae and to help the robot “make decisions” about the best way forward.
The future is here
All the robot research groups that have been working on insect-inspired robots such as Genghis, RHex, Sprawl and Whegs, and the students that came out of these laboratories to start their own groups, have branched out into other areas of research (some of which involve insects). The focus may have shifted to:
Locomotion beyond walking and running.
Rapid inversion like cockroaches and geckos do to get under ledges.
The biggest challenge to robotics is powering small insect-sized robots. We can still learn a lot from insect’s operational duration. Making a robot work for 5 min is great. However, insects work for days on end. We need to incorporate similar power management strategies and power budgets into our robots. This becomes even more critical as we scale down in size because available power doesn’t scale linearly with length. Less power can be stored per unit volume as you get smaller because the power/packaging ratio goes down. The miniaturization and power issue is especially critical for developing smaller flying robots. (Insect flight will be covered in a later blog post)
One area where we can also still learn from insects, and which in my mind has been somewhat ignored, is the fact that insects can recycle large parts of their exoskeleton...maybe this can become a focus too. I will explain more about the beauty of the materials that make up the insects exoskeleton in one of my next few blog posts.
Until then, all you need to remember about this post is that “The truth is out there”.
To some of my entomology friends the title of this blog may not seem particularly original. That is probably because they are familiar with the book “Insects Did It First” by Roger D. Akre, Gregory S. Paulson and E. Paul Catts (1992). I had my heart set on this blog title (with the subtitle “Can Engineers Do It Better?”) before I was aware of the book.
My copy of the book with the same name as this blog. (Picture by Marianne Alleyne)
All three authors were entomologists and associated with Washington State University (Dr. Paulson now teaches at Schippenburg University of Pennsylvania). The book “Insects Did It First” is a collection of ideas, started in 1964 by Akre, that linked an “advanced” human technology to insects. The book is a perfect example of how to get the general public to become more interested in the natural history of insects. The book is even more endearing because of the wonderful, often humorous, drawings by Catts.
Typically whimsical drawing by E. Paul Catts from “Insects Did It First”. – picture featured on Gregory Paulson’s website (click drawing).
All 81 short “chapters” of the book cover an achievement in which insects were far ahead of humans. Some examples are obvious and famous (e.g. insects as builders of energy efficient structures), other are less well known to non-entomologists (e.g. preserving and storing food without freezing).
In some ways this blog is similar to the Akre, Paulson and Catts book – but using a media that may be more accessible to more people. Just like the author-trio my ultimate goal is to promote insects as inspirational to those outside of entomology. I hope to especially reach engineers, designers and entrepreneurs. I may cover some of the same topics, but since the book was last published in 1992 (Dr. Akre passed away in 1994 and Dr. Catts in 1996) I will be able to give more updated information. The blog will also be different in that I want to go beyond natural history and delve a little bit deeper into the topics of technology and innovation. In addition, there are characteristics of insects bodies, their behaviors, the ecosystems they live in, etc. that I think have not yet been considered in depth by engineers. I will promote those topics too. For instance, the Akre book does not cover the springing mechanism of Collembola which I covered in my previous post (maybe because Collembola are not insects?).
Ultimately I hope that my blog will be thought of as fondly as the Akre book.
(Stay tuned for next week’s blog post (also on a topic not covered by the Akre book) on how insect-inspired robots evolved between famous X-files episode and now.)
And here it is. Behold the best blog-banner ever – created by Nils Cordes*!
Of course, the premiere of such a great banner also requires a blog post that explains it. So let me try.
The animal featured in this blog’s banner is a springtail from the hexapod lineage Collembola. Collembola are not insects but entomologists are an inclusive bunch so we gladly incorporate spiders and entognathous creatures into our studies and teachings.
Springtails are very likely the most abundant arthropods on earth. They occur in the soil (different species at different depths), in leaf litter, moss, under logs, etc. One of the most distinguishing features, if you can consider anything on an animal that is only 0.12 to 17 mm long distinguishing, is the forked furca at the posterior end of the animal. The furca is present in a lot of species, but not all. Those that live deeper in the soil usually lack the structure because they do not need it since its main function is for jumping.
Generalized “elongate” (top) and “globular” (bottom) Collembola. Furca (springing mechanism) in red – the springtail at the top has the mechanism partly retracted and the springtail in the bottom picture has the furca extended. (Marianne Alleyne)
Collembola species can have varying body shapes, but generally there are those with elongated bodies and those with more globular bodies. Collembola can walk, run and climb, but the locomotory specialty that they are best known for (and which seems to be rather ancestral) is jumping.
Globular springtail Dicyrtomina saundersi. Body length = 1.7mm. Picture by Lord V. (Used with permission.)
Elongate springtail. Body length = 2.3 mm. Picture by Lord V. (Used with permission.)
This excellent picture is of a springtail’s underbelly. The picture was taken by Lord V as a springtail walked over a glass slide. Clearly visible is the forked furca that can propel the animal into the air. (Used with permission.)
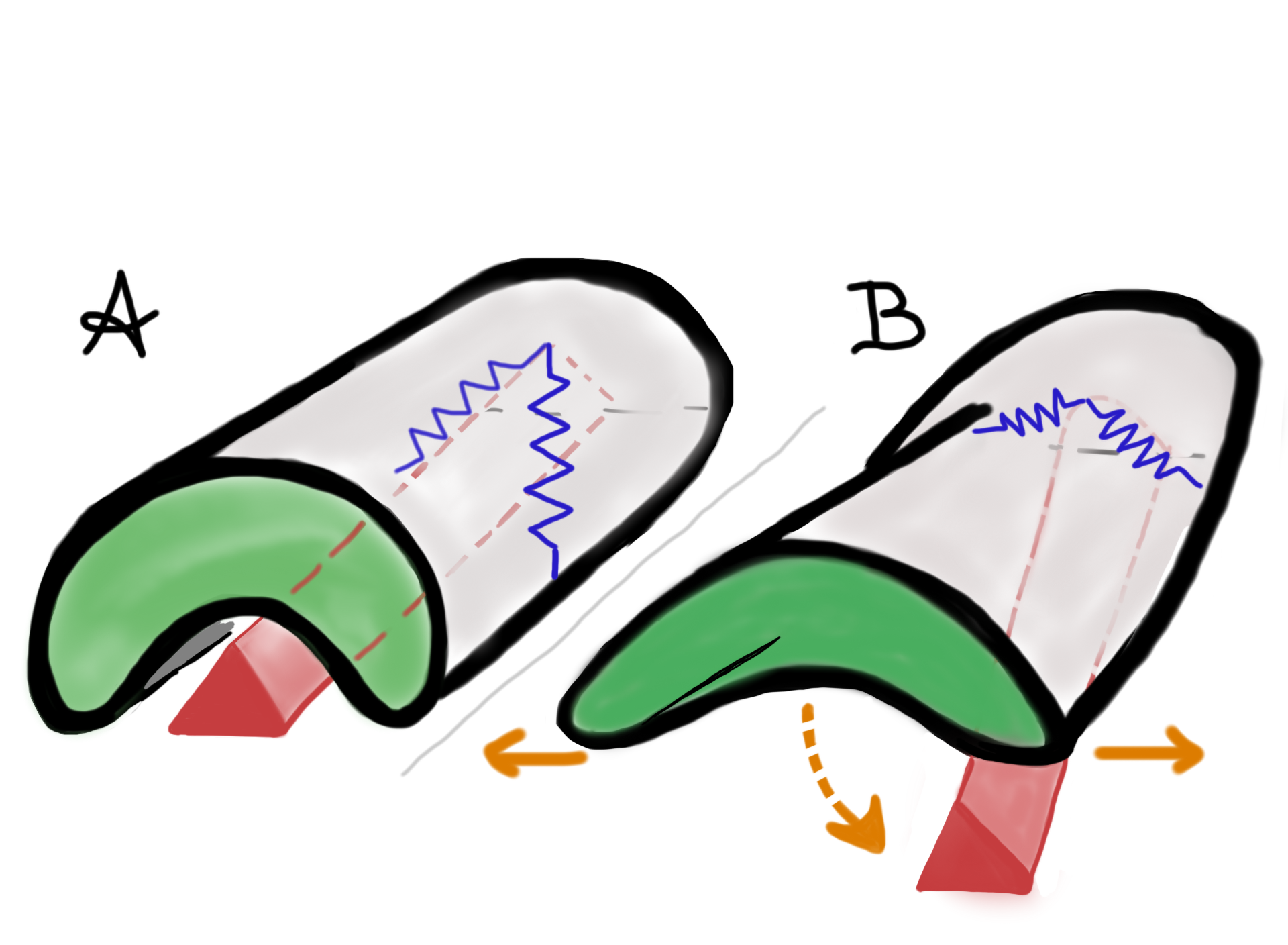
Collembola can jump multiple times in a row, those with globular bodies and more advanced tracheal systems more often (1). In general, springtails tire easily so that jumping is usually only used as an escape mechanism. The jump can take the animal in any direction. Since the furca is located at the extremity of the body, directly beneath the center of gravity, the dynamics of the jump cause the body to rotate head over end. Some Collembola species can jump very high, others take a shallower trajectory but land far away from point of takeoff.
This jumping escape response is quite successful but it does require modifications of the entire body plan. The cuticle, the (hydrostatic) endoskeleton, tendons, and muscles all work together to manipulate the body in such a way that the propulsion is optimal. How exactly this happens is not very well understood, yet this system holds inspirational lessons for passively compliant locomotory structures.
At rest the furca is held within a ventral groove of the abdomen. At the time of the jump the furca moves from this resting (retracted) position to the extended position. Based on morphological and kinematic observations (there is no direct experimental evidence) it appears that as the furca moves it compresses a “spring”. After it passes a critical point of extension the spring releases all the energy, which in turn causes the springing organ to snap out at high speed. If this happens as the springing organ hits a substrate a force is created that propels the animal upward.
The springing mechanism of a generalized springtail; partially retracted (left) and extended (right). (Marianne Alleyne)
What exactly comprises this “spring” is not clear. Earliest experiments done by Manton (2) in the early 1970s concluded that to evert the springing organ the body’s hydraulics (pressure on the fluid that makes up most of inside of the body = hemocoel) was important. However, later in the 1970s, Christian (3) concluded that direct muscle action, and not necessarily hydraulics, was the main force inducer. In the 1990s, when high-speed photography had advanced greatly, Brackenbury and Hunt (4) concluded from their experiments that hydraulic forces created by pressurizing the hemocoel increases tension on abdominal sclerites (the exoskeletal plates) that results in a click mechanism that propels the animal into the air. All these studies do agree that elastic elements within the base of the springing organ and within the exoskeleton, as well as the body as whole, are important too. To what extent is not known.
Click mechanism model of the furca (red) and distal end of abdomen. The furca, at rest, is retracted into an abdominal ventral groove. A pair of “basal rods” (springs, in blue) are embedded in ventral and lateral parts of the abdominal sclerites 4 & 5, these springs also attach to the apex of the furca. The spring/click mechanism gets help from muscle and active dorsiflexion of the body (in orange), to release the spring organ from the groove. After the furca passes a critical point of extension the spring releases all the energy. (Drawing by Marianne Alleyne based on Brackenbury & Hunt, 1993)
Imagine a beam or a chopstick that’s flexible transversally but somewhat stiff longitudinally. If you compress it, it doesn’t change…up to a point. Then it ‘snaps’ out and buckles. You get a rapid displacement as all the strain energy is released. The exoskeleton of the springtail does a similar thing. It stores the strain energy and then goes through a snap-through buckling phenomenon to produce large strain motion which is then amplified by the tail and presto…springtail in motion.
Many insects, and other animals, use musculoskeletal springs that are incorporated into the complete body plan. These springs help achieve a high rate of acceleration, or a further jumping distance, and help save metabolic energy. Based on these findings compliant structures and materials have been incorporated into bioinspired legged robots (5). Compliant legged robots achieve a few important things: increased energy efficiency, increased speed, ability to avoid obstacles (in case of jumping robots), and the ability to use more simplified controls to enable enhanced gait control and shock absorption. Springs in bioinspired robots have used elements such as airsprings (e.g. compressed air) and compliant materials, but improvement is still possible. Airsprings, for instance, are not very efficient because they end up converting much of the energy they store into heat. In addition, some of the compliant materials are better than others. Rubbery materials, like elastomers, tend to have a fair bit of viscosity in them and so some (maybe lots) of the energy that it stores is lost to heat as well. For high efficiency, most robotic-type systems currently use mechanical springs (i.e. metals). Bioinspired robots also incorporate series elastic actuators that have linear springs intentionally placed in series between the motor and actuator output, which results in the actuator being bulky.
The variety of jumping mechanisms among insects is great (think: click beetle, flea, grasshoppers, treehoppers, etc.). The intriguing aspect of the jumping mechanism in springtails is that it operates so efficiently at a very small scale, much smaller than any bioinspired robot that has been developed. In the future we will be able to manufacture almost microscopic devices incorporating different characteristics into small structures using “springs” and compliant materials.
Maybe we can incorporate locomotory mechanisms that propel the object, using very little energy. Inspiration for what materials to use and how to construct the object can be found through further study of the springtail’s click mechanism. Somewhat surprisingly not much research has been published on this system since the 1990s. Yet with help from today’s high-speed cameras and microscopy techniques we should be better able to understand how the springtail propels itself. Advanced computer aided engineering (CAE) tools, like finite element analysis (FEA), could be used to augment the visual data and elicit some fundamental internal characteristics that are not visibly detectable.
By researching this topic I thought of a few applications for technologies based on the Collembola’s spring mechanisms. Click mechanisms at the scale of a springtail’s springing mechanism could possibly aid stent design or inspire development of other deployable structures that snap open or closed based on certain environmental conditions. Maybe small springing mechanisms can be incorporated in groups and serve as strain sensors on bigger structures. And who wouldn’t welcome millimeter-sized robots that can perform in a futuristic “flea circus”?
PLEASE LEAVE A COMMENT (OR CONTACT ME VIA TWITTER @COTESIA1) IF YOU HAVE ANY NOVEL IDEAS FOR A SPRINGTAIL INSPIRED JUMPING MECHANISM. JUST IMAGINE.
REFERENCES:
(1) B. Ruhfus and D. Zinkler, Investigations on the sources utilized for the energy supply fueling the jump of springtails, Journal of Insect Physiology, Volume 41, Issue 4, April 1995, Pages 297-301, ISSN 0022-1910, 10.1016/0022-1910(94)00122-W.
(2) S. M. Manton. The Arthropoda: Habits, functional morphology, and evolution. Clarendon Press, Oxford, 1977. ISBN: 019857391X
(3) E. Christian. The jump of the springtails. Naturwissenschaften, Volume 65, Issue 9, 1978, Pages 495-496, 10.1007/BF00702849
(4) J. Brackenbury and H. Hunt. Jumping in springtails: mechanism and dynamics. Journal of Zoology, Volume 229, Issue 2, 1993, Pages 217-236, ISSN 1469-7998, 10.1111/j.1469-7998.1993.tb02632.x
(5) Z. Zhou and S. Bi. A survey of bio-inspired compliant legged robot designs. Bioinspiration and Biomimetics,Volume 7, Issue 4, 2012, 20 pages 10.1088/1748-3182/7/4/041001
*My friends can attest to the fact that I have been talking for a long, long time about starting a blog about how we can use insects to inspire new technologies. One of these friends who had to humor me for so long is Nils Cordes. I met Nils when he was a student at Illinois, but he is currently finishing up his PhD at the University of Bielefeld in Germany. Nils is a great scientist, and a great communicator. He is also a wonderful artist. He offered, those many years ago, to create some art work for this (then still imaginary) blog that I was going to use to communicate my love of insects. And he did…behold the best blog banner EVAH!
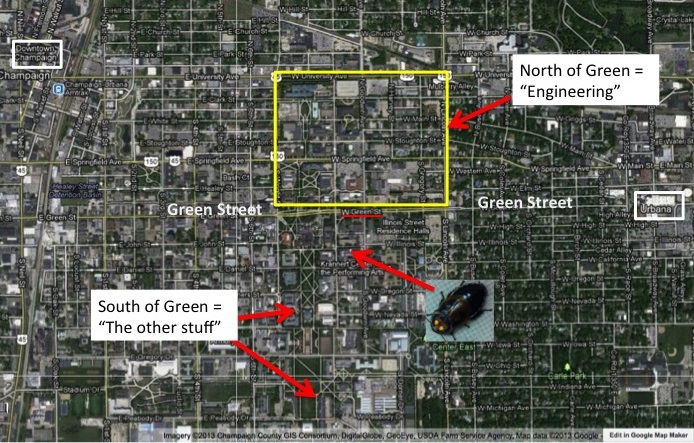
People at the University of Illinois at Urbana-Champaign often make a distinction between “Those North of Green Street” and “Those South of Green Street”. In a town that is almost perfectly arranged as a square grid, Green Street runs through the campus, connecting downtown Urbana to Campus town and to downtown Champaign.
Google Map of the UIUC campus
Located North of Green Street are almost all buildings associated with the College of Engineering and South of Green Street is everything else (it seems). North of Green is where the money is, South of Green not so much (it might appear to an Entomologist). South of Green is where the slackers roam, and North of Green where the studious students are holed up (it might seems to the average Engineer).
This (for the record: incorrect) image is not really helped by the fact that Green Street is quite a dangerous street to cross. Unless you have courses on the other side, why would you risk your life? Quite a few students rarely have to make this choice during their 4+ years on campus. However, I try to do my part to make students experience life on the other side of Green Street. Students from different disciplines benefit from interactions. And, Hello!, the same goes for faculty and staff.
Last week I made students risk their lives and cross Green Street, just so I could change their future by having them touch some cockroaches. I am happy to report that all students (mostly engineers from North of Green), who are enrolled in the ENG333 course, arrived safely at my lab (located South of Green), and that there might be at least a few students out of about 40 who enjoyed the experience.
This semester the displays were kind of heavy on the cockroaches because of the 15 or so different species that would make an appearance at the 30th Insect Fear Film Festival to be held a few days later. But we also had live ants, flies, lubber grasshoppers, etc.
The goal of the “field trip to the Alleyne lab” is to get students to look at insects in a way they have never looked at, or touched, them before. I try not to talk for the full 45 minutes, I encourage students to ask questions and thus to guide the discussion, but I do some talking and I try to cover at least 2 or 3 of the 6 topics outlined below. In the future I will cover these topics in more detail here on the blog. I leave this detail out for this field trip since I want the students to be guiding the creative process (remember, the course is called Creativity, Innovation and Vision).
1. Structural Color in Insects
Butterfly – nanostructures -> structural color display
Many butterflies and beetles have nanostructures that give their wings and bodies iridescent structural colors. I made this Prezi presentation a few years ago. The information is basically what I explained to the students visiting the lab.
2. Cockroach-inspired robots
Bioinspired robots have become quite popular over the past few decades. The most famous examples are the robots that were inspired by cockroaches (again, I will blog more about this in the future). Engineers realized that the stability of the roaches due to their tripod gait, and their ability to go rather fast over many different types of terrains were all characteristics that would serve robots well.
This short video shows both the tripod gait, the stability and the quickness of the Madagascar hissing cockroaches (Gromphadorhina portentosa). (The tweet itself shows one should not tweet while exhausted – I couldn’t type strate 😉 )
I also pointed out the many “simple” yet elegant sensors that insects use to get around in their environment, and to taste stuff, and to find each other. This is easy to see in the cockroaches (see video in previous section) as they try to navigate obstacles – they use their antennae, for instance. I also encouraged the students to hold a lubber grasshopper and to point out some of the sensors that are easily visible (compound eyes, antennae, sensors on mouthparts), and then told them of the ones that are not so obvious (mechanoreceptors, for instance).
Gwyn and I discussed the live Manduca sexta (tomato hornworms) and pointed out the different types of cuticle an individual makes during its lifetime. The cuticle is made up primarily of chitin and protein, is build “from the ground up” in an hierarchical manner, and large parts of it get recycled at each molt. In the picture below the caterpillar on the left will initiate pupation in a few days and then eventually turn into a pupa like the one on the right. The caterpillar’s cuticle is soft and bendable, the pupa’s is hard and tough.
Manduca sexta (Tomato hornworm) caterpillar (L) and pupa (R)
5. Social Insects
There is always so much to discuss when it comes to social insects, so we usually have the students just ask questions. In the past I have shown students leaf-cutter ants.
Leaf-cutter ant colony observed by ENG333 students (in 2011). It is always interesting to watch emergent behavior get things accomplished – in this case getting leaf material from the tray in the foreground into the nest located in the tray which is in the background.
This year Adrian told the students about a local carpenter ant species, Camponotus pennsylvanicus, and (my favorite) trap-jaw ants. This semester the trap-jaw ant species was Odontomachus rixosus, a species collected in Cambodia.
Harvester ant colony (two arenas in the foreground) and Trap-jaw ant colony (rectangular arena in the background
As a bonus, Adrian also brought in a zinc nest cast of Messor pergandei (an harvester ant from Arizona).
Zinc cast of an harvester ant species, Messor pergandei, that always makes nests that run down at an angle. Most other species have nests that go straight down. This only about 1/2 of the complete size of the nest.
Of course, we always have to show the videos of trap-jaw ants jumping “with their mouth”, and then crashing back to earth – and surviving. I tried to make the students think about the amount of force a small animal can generate with the right muscle and cuticle, and that it is amazing that the cuticle does not shatter upon impact.
6. Other insects
Other live insects that made an appearance were:
The milkweed bug, Oncopeltusfasciatus
Mealworm larvae, Tenebrio sp.
House flies, Musca domestica
Death-feigning beetles, genus Cryptoglossa
Lots of different species of cockroaches, including:
The cutest cockroach
Lucihormetica verrucosa, warty glow spot. This individual was given the name Frodo.
And the one from X-files fame, Blaberus giganteus, the giant cave cockroach (see next blog post for more details about X-files and their roaches)
Insect nervous system dissection slides (cockroach)
Slides of cockroach nervous systems.
Gwyn talking Science, and student scribing Science!
Some things I would like to do differently next time.
1. Prepare some questions that students have to fill out before they come on the field trip. I would ask them questions about what they know about biomimicry and bioinspiration, and about the videos they were supposed to watch.
2. While in the lab some students took to the project/process easily and did exactly what I wanted them to do: ask questions, take notes, make sketches. To help others get “jump-started” I might also come up with a worksheet that encourages students to Science Scribe the whole adventure. This may require clipboards and thus a trip to to the office supply store. YEAH!
Adrian talking to one group of students about ants.
All and all, it was a fun afternoon. I think we made a few students think about:
Insects, and other animals and even some plants, can serve as inspiration for technological innovation.
There is actually some really cool stuff going on South of Green. Some of it may even be worth a trip across Green Street.
At this point in the semester I introduce myself to the students in the Engineering courses with the title Creativity, Innovation and Vision. There is both an undergraduate (ENG333) and a graduate version of this course (ENG598).
The originator of the course is Dr. Bruce Litchfield. Students in these courses learn that their own state of creativity is not as static as they might expect. Bruce and his collaborators also do research on creativity enhancement; paying special attention to the ways in which engineering students currently incorporate creativity, since it has been shown that for engineering students creativity does not increase as they move through their college courses (the same is likely to be true for students in other disciplines).(1)
“Personal creativity enhancement via exploration of the nature of creativity, how creativity works, and how to envision what others may not. Practice of techniques and processes to enhance personal and group creativity and to nurture a creative lifestyle. Application to a major term project providing the opportunity to move an idea, product, process or service from vision to reality.” (2)
The courses are quite popular with students from all over campus, not just Engineering. Many of the students who take the graduate level course become teaching assistants for the undergraduate course in subsequent semesters. (I think in this case the term ‘facilitators’ instead of TA is more applicable)
BioInspiration (formerly BioCreativity)
Over the last ~3 years I have worked with Bruce and the TAs on a module we call BioCreativity BioI. It is basically a module on BioInspiration or Biomimicry. I now kind of regret coming up with yet another term for a field of study that suffers from much confusion due to terminology already, but students seem to like the title because it fits into the focus of the course so perfectly. However, if we adhered to proper terminology more rigorously it should be acknowledged that BioCreativity is actually the combination of biology and art, not biology and technology, as we use it here. [Note: in 2014 we realized that the term Biocreativity created too much confusion and we decided to name this module BioI or BioInspiration].
The BioInspiration module is divided into four class meetings and each meeting is separated by 2 or 3 weeks [Note: in 2014 we also decide to condense the module since students felt they were not able to focus on this one task if they had all these other topics being thrown at them too.] This week I met the students of two ENG333 sections. This semester a large majority of students are engineers (mechanical, chemical, civil, electrical, bioengineering). A number of students are computer science majors, and advertising majors. A couple of students are majoring in the arts, such as creative writing and graphic design. Students from the humanities are also represented, by majors in philosophy and anthropology. In other words, it is quite a diverse group of students eager to learn how to enhance their creativity.
During our first meeting this past week I introduced the students to the topic of Biocreativity. I mostly talked unscripted, but I also had a pretty PowerPoint behind me with amazing pictures by Alex Wild (http://www.alexanderwild.com/). [Note: in 2014 the course will have 7 or more sections. Too many for me to visit. We have decided to therefore put this first lecture on video which will be presented to the students during the class.]
I continued the introduction by explaining how I became interested in Bioinspiration. I like to tell the students that it is all my husband’s fault. I am married to a mechanical engineer and over the 25 years that we have known each other, we have taken many a road trip. Usually during these trips we end up “discussing” why insects are better/worse at “doing stuff” than human engineers. In the beginning (the first 24 years) he always ended the argument by saying something like: “Well, sure that might be a cool thing that insects can do, but can they fly 500 people across an ocean? No? Well, there then!” My interest in teaching modules, courses, and now this blog on Bioinspiration is all because I really want to learn how to win this argument.
“Biomimicry Shoe” by Marieka Ratsma and Kostika Spaho. Interesting, yes. Pretty, maybe. Biomimicry, definitely not. Photograph by Thomas van Schaik.
I then very briefly explained what I mean by Biomimicry and Bioinspiration. I do this quickly because the topic of definitions might evaporate all creativity out of these students. I put up Janine Benyus’ (Biomimicry3.8) Life’s principles, and also Robert J. Full’s quote about evolution working on the just good enough principle. I actually spend more time on what I think biomimicry and bioinspiration is not. Students see these types of examples often in popular media because the terms have become buzzwords.
Why have biomimicry and bioinspiration become buzzwords? In my opinion it is probably because people like to think that if we copy/mimic/emulate nature, or at least base some or our new engineering designs on nature, then it is probably also more sustainable. And sustainability is itself a buzzword. I stressed in my presentation that that is not necessarily the case. The most famous example of bioinspiration is probably Velcro, which is made from synthetic materials that are not biodegradable and cost a lot of energy to produce. For many scientists who are inspired by nature and use biomimicry or bioinspiration as a guide it is not sustainability per se that drives them. It is a guide to making new basic biological discoveries, or to innovate and solve a technological problem. “Why does an animal or a plant do that? And how can we use that what I have learned in a new technology?”
Next I make a very controversial statement: “I think my husband is basically correct.” Of course, nature has not been able to carry 500 people across an ocean. Primarily because of the issue of scale. Nature works at a much smaller scale than we humans usually do. However, we currently live during very exciting times, where we can find inspiration for innovation at a smaller scale. We can now image at the nano-scale. That means that we can see structures and processes at a scale where very important things in nature happen. At the same time we are starting to be able to manufacture at that size scale too. We can start to build structures the way that nature builds materials and structures; hierarchical and from the bottom up.
Just consider an ant. Think of the interesting aspects of an ant’s body and life history. All these apsects have the potential to inspire us. (These are subjects I will blog about in greater detail at a later point).
Exoskeleton (cuticle). Multifunctional. Made from relatively few elements (compared to all the elements from the periodic table we use to manufacture our multifunctional materials). One individual often has cuticle that has different characteristics – soft (larva, abdomen) or hard (adult, head), for instance. And on top of that, when molting occurs in the larval stages most of this cuticle is recycled and used in the new cuticle. No toxic substances required. All of life’s principles satisfied.
Located on the surface of the cuticle are nanostructures that can help capture moisture, or give an insect color (as is the case in the Morpho butterfly).
The locomotory mechanisms of insects, including ants, has inspired many bioinspired robots. I have tried to keep up with all the different bioinspired robots on this Pinterest Board.
Insects, even tiny ones like this ant, have many interesting sensors on their bodies: compound eyes, simple eyes, antennae, mechanoreceptors, etc.
Ant and termite nests have also been of interest for bioinspired architecture since through cooperative behavior they can build structures that are relatively stable and require few inputs (Again, unlike our own structures).
And sociality in ants, the cohesion that exists between these “small brained” insects, has inspired electrical and computer engineers.
And so on.
These are all examples of inspiration points from just an ant.
By this point it was my hope that students understand the possibilities that exist. I gave them some tips on how they can become “bioinspired”.
Avenues to becoming BioInspired (as a student in CIV)
1. Delve into biomimicry and bioinspiration basics
Students were asked watch two videos before the next BioCreativity meeting.
Look at things from different, less familiar angles. Look at a whole tree (Why is a tree that shape?), go closer (Why is the bark textured like that?), go even closer (Why does moss grow in those crevices).
Sketch or take pictures
Bring your friends – talk about what you are seeing.
Biomimetics in Architecture: Architecture of Life and Buildings. Petra Gruber. 2010
Biomimicry: Innovation inspired by nature. Janine M. Benyus. 2002
The smart swarm: How to work efficiently, communicate effectively, and make better decisions using the secrets of flocks, schools, and colonies. Peter Miller. 2010
Learning from the octopus: How secrets from nature can help us fight terrorist attacks, natural disasters and disease. Rafe Sagarin. 2012
Darwin’s devices: What evolving robots can teach us about the history of life and the future of technology. John Long. 2012.
How to catch a robot rat: When biology inspires innovation. Agnes Guillot and Jean-Arcady Meyer. 2010.
Etc.
Use Social media
For example Twitter. I suggest you follow these folks because they often tweet links to interesting bioinspiration or biomimicry (and thus biocreativity) topics.
And then I sent the students off into the world, to get inspired. Actually, I explained a little bit more about the project we want them to do, but I will leave those details until the next blog post about BioCreativity.
(1) This research by Burgon, et al. (under review) measured the creativity of first- and fourth-year engineering students using two nationally-normed creativity assessment instruments. I will blog more about this work when it has been published.
(2) More information about the Creativity, Innovation and Vision courses:
Two videos that introduce the topics discussed in the courses can be seen here:






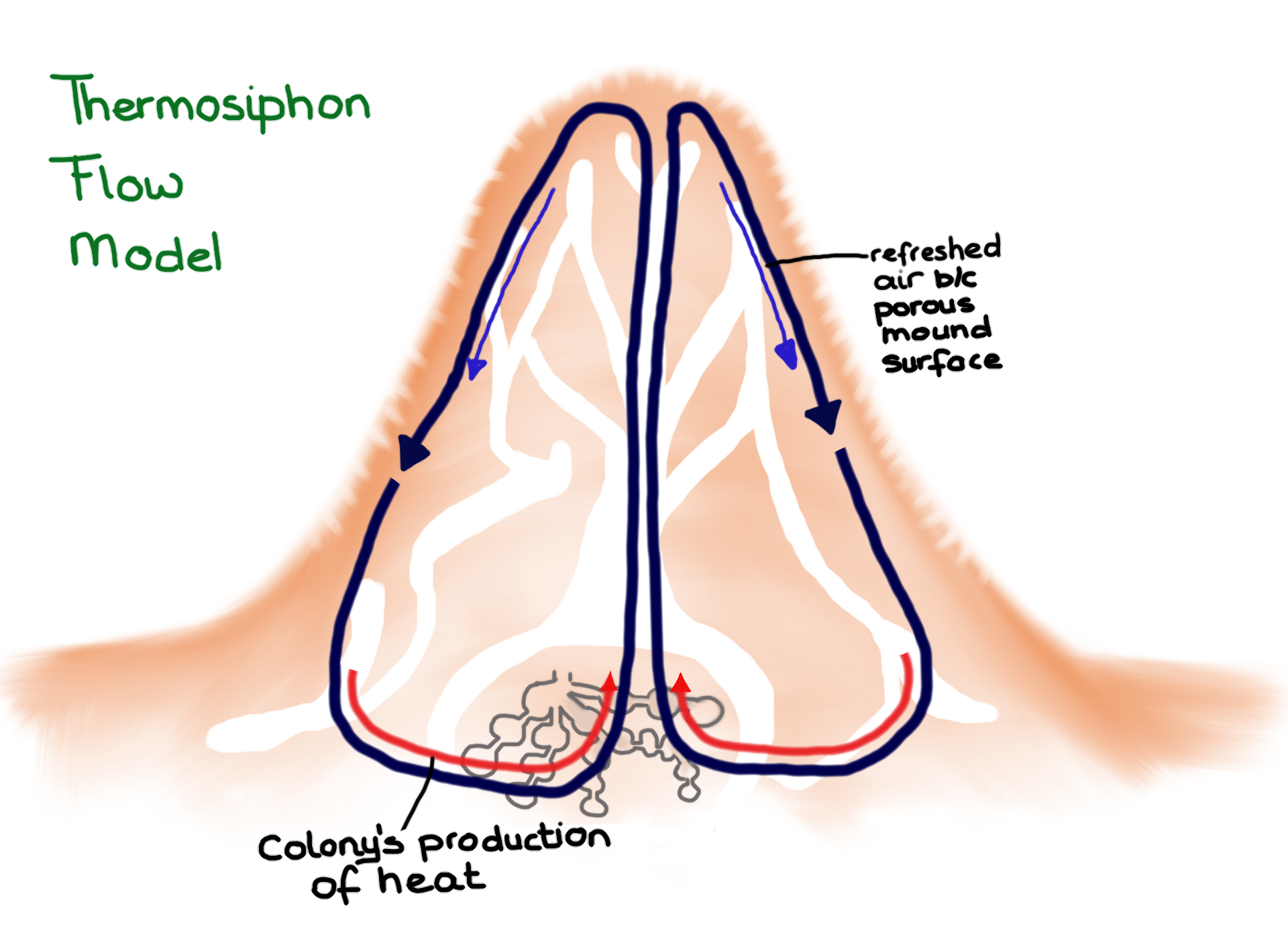


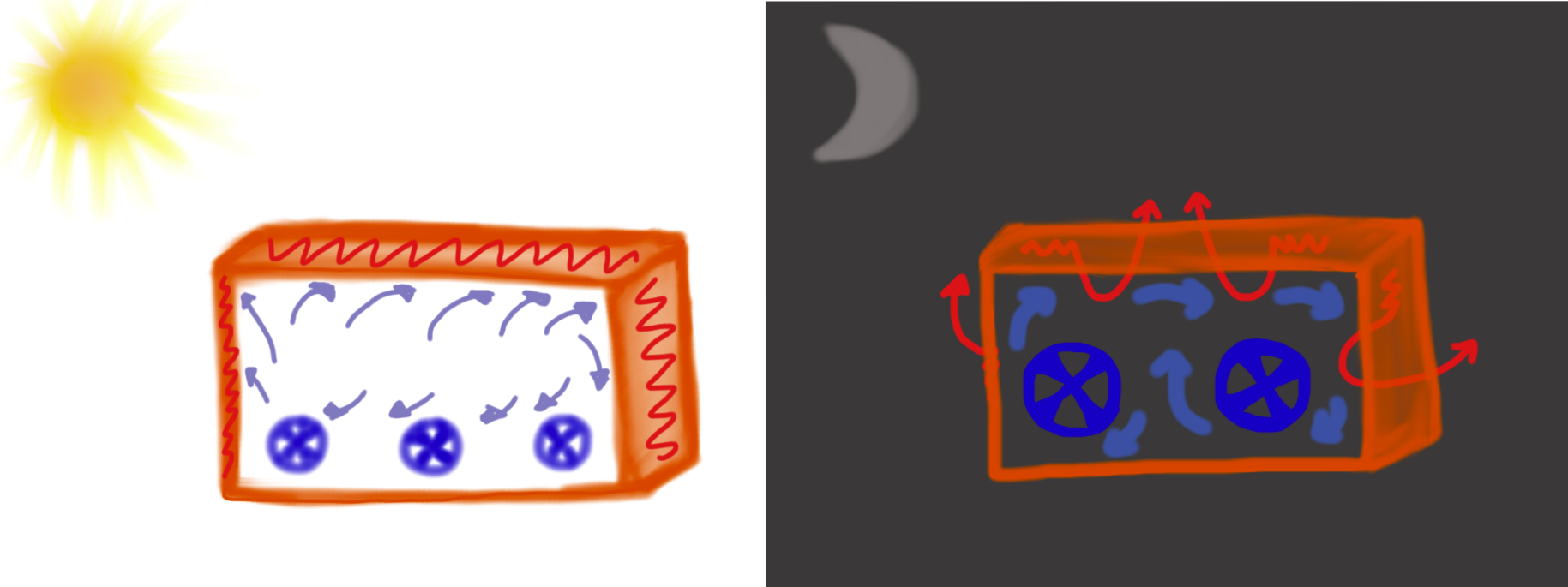
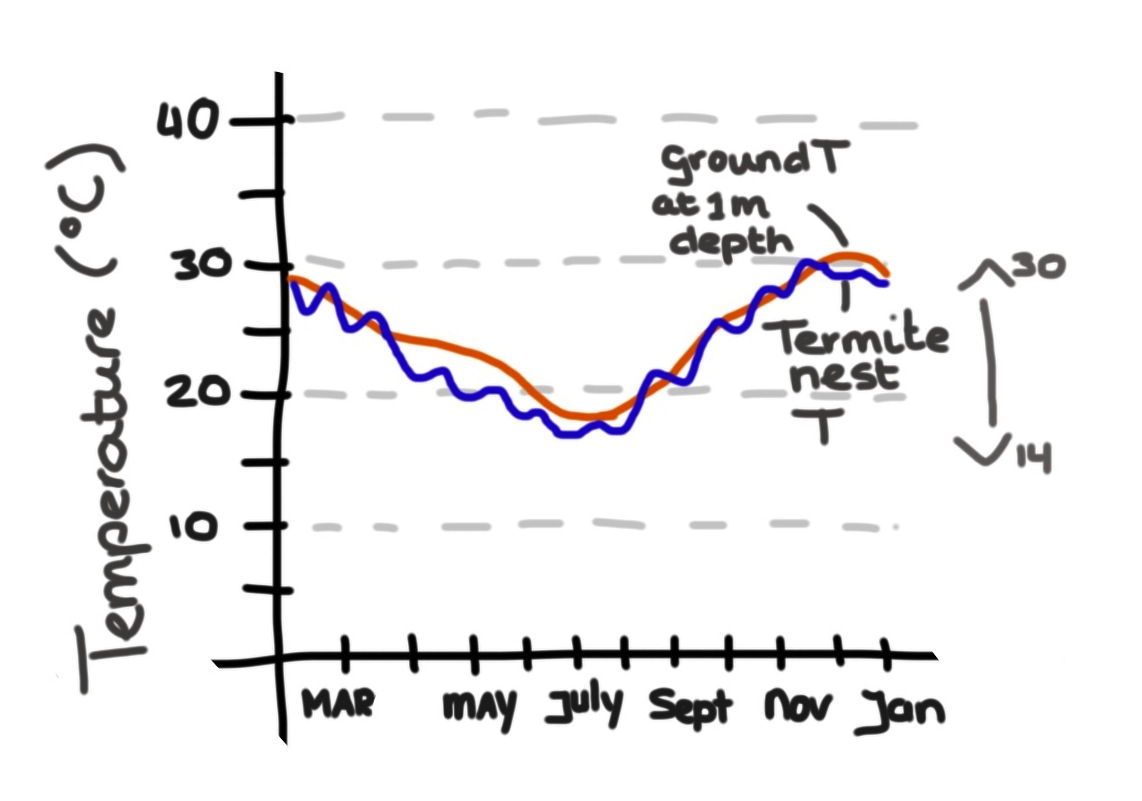









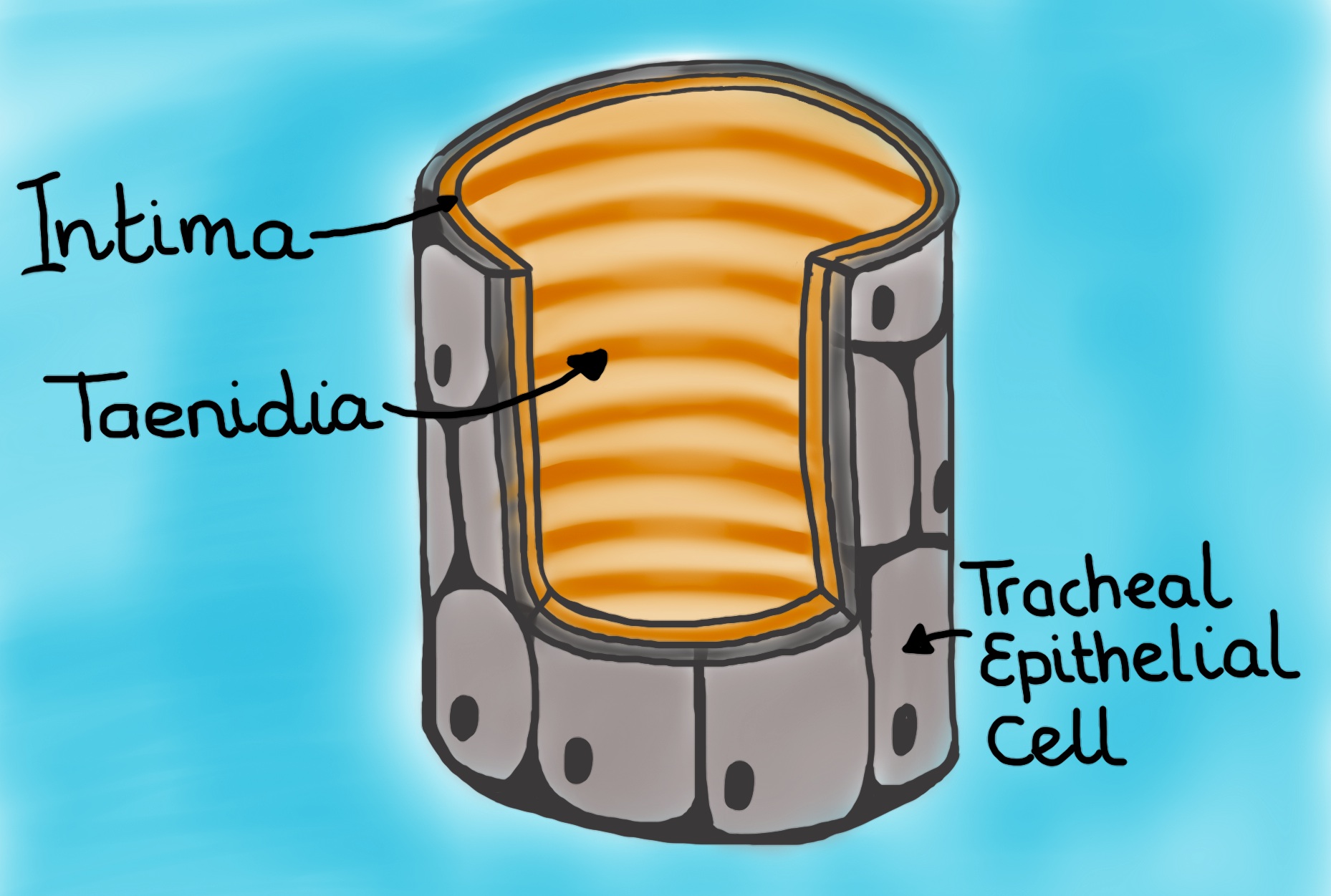




")